
Лекция для врачей "Ультразвуковая анатомия щитовидной железы". Лекцию для врачей проводит профессор В. А. Изранов.
Дополнительный материал
Анатомия щитовидной железы
Щитовидная железа (ЩЖ) является самой крупной эндокринной железой человеческого организма, имеющей только внутрисекреторную функцию. Ее масса у взрослого человека составляет около 15-20 г. ЩЖ состоит из двух долей и перешейка, располагающихся на передней поверхности трахеи и по ее бокам (рис. 3.1). Иногда от перешейка отходит дополнительная пирамидальная доля. ЩЖ развивается из выпячивания середины дна первичной глотки. Ее закладка происходит на 15 неделе внутриутробного развития, к 18-20 неделе она начинает продуцировать тиреоидные гормоны. При нарушениях эмбриогенеза ЩЖ могут развиваться различные аномалии ее расположения. К ним относятся кисты щитоязычного протока, язычная ЩЖ, а также срединные и боковые остатки тиреоидной ткани.
ЩЖ состоит из клеток двух разных видов: фолликулярных и парафолликулярных (С-клетки). Фолликулярные клетки, продуцирующие тироксин (Т4) и трийодтиронин (Т3), формируют в железе многочисленные фолликулы, каждый из которых состоит из центральной полости, заполненной коллоидом, главной составляющей которого является белок тиреоглобулин, окруженной одним слоем кубовидных эпителиальных клеток (рис. 3.2). Парафолликулярные клетки (С- клетки) продуцируют белковый гормон кальцитонин.
Т4 и Т3 синтезируются из аминокислоты тирозина. Кроме того, существует биологически неактивный реверсивный трийодтиронин (рТ3), который образуется при дейодировании внутреннего кольца Т4 (рис. 3.3).
Необходимым структурным компонентом тиреоидных гормонов является йод. Этот микроэлемент практически полностью всасывается в кишечнике, откуда в ионизированной форме поступает в плазму крови. Перенос йода внутрь тироцита энергозависим и осуществляется сопряженно с обратной транспортировкой натрия натрий-йодидным симпортером (NIS). В клетках ион йода окисляется, после чего подвергается органификации, присоединяясь к остаткам молекул аминокислоты тирозина, которые присутствуют в тиреоглобулине, синтезируемом фолликулярными клетками. После этого тиреоглобулин изменяет свою структуру таким образом, что тирозильные остатки сближаются друг с другом, облегчая реакцию конденсации между ними. Йодирование тирозильных остатков и реакция конденсации происходят при участии тиреоидной пероксидазы (ТПО), локализованной в микросомальной фракции фолликулярных клеток. Далее содержащий связанные йодтиронины тиреоглобулин транспортируется через апикальную мембрану и откладывается в коллоидной полости фолликула, после чего вновь поступает в тироциты, перемещается к их базальной мембране, высвобождает йодтиронины, которые выделяются в кровь. Попав в кровь, Т4 и Т3 разносятся по организму в основном в связанном с белками плазмы (тироксинсвязывающий глобулин, преальбумин (транстиретин), альбумин) виде. Период полужизни в крови для Т4 равен 7-9 дням, для Т3 - 2 дням.

Рис. 3.1. Щитовидная железа

Рис. 3.2. Гистологическое строение щитовидной железы
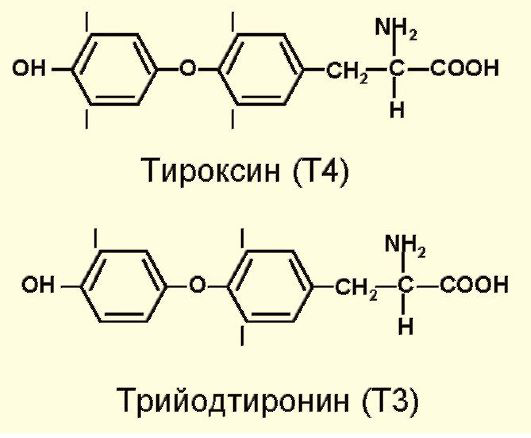
Рис. 3.3. Структура гормонов щитовидной железы
Свободные йодтиронины относительно легко проникают сквозь мембрану клеток. Внутриклеточные эффекты тиреоидных гормонов тесно связаны с процессами их метаболизма (в первую очередь с механизмами дейодирования). Самым важным из таких превращений является конверсия Т4 в более активный Т3 . Поскольку с рецепторами тиреоидных гормонов непосредственно взаимодействует преимущественно Т3 , а не Т4, последний принято рассматривать как прогормон, а Т3 - как истинный гормон. Т3 , воздействуя на специфические ядерные рецепторы, которые присутствуют в большинстве клеток, приводит к изменению экспрессии различных генов, что проявляется физиологическими эффектами тиреоидных гормонов, главным из которых является регуляция и поддержание основного обмена. Лишь 5- 10 % циркулирующего в крови Т3 синтезируется непосредственно ЩЖ; его большая часть образуется в результате дейодирования Т4 в периферических тканях. Превращение (конверсия) Т4 в Т3 катализируется различными дейодиназами, обладающими тканевой специфичностью.
Функция ЩЖ регулируется аденогипофизом по принципу отрицательной обратной связи. Тиреотропный гормон (ТТГ) гипофиза стимулирует продукцию тиреоидных гормонов, которые подавляют продукцию ТТГ (рис. 3.4).
Эффекты тиреоидных гормонов многообразны. Они обеспечивают поддержание основного обмена в большинстве клеток, регулируя их метаболическую активность, а также процессы пролиферации и апоптоза. Нормальный уровень тиреоидных гормонов необходим для функционирования всех без исключения систем организма, а при нарушениях функции ЩЖ патологические изменения носят полисистемный характер. Тиреоидные гормоны необходимы для формирования нервной и других систем плода и новорожденного. Дефицит тироксина в этот период приводит к значительным неврологическим нарушениям. В целом эффект тиреоидных гормонов традиционно описывается как калоригенный: под их действием происходит повышение основного обмена за счет роста потребления кислорода и увеличения теплопродукции тканей.

Рис. 3.4. Регуляция секреции тиреоидных гормонов




")
")
")
")
 (тип с усилением стромального компонента)")
 (тип с усилением стромального компонента)")





 зоба")
 зоба")
 зоба")
 зоба")
 зоба")
 зоба. ЦДК")




